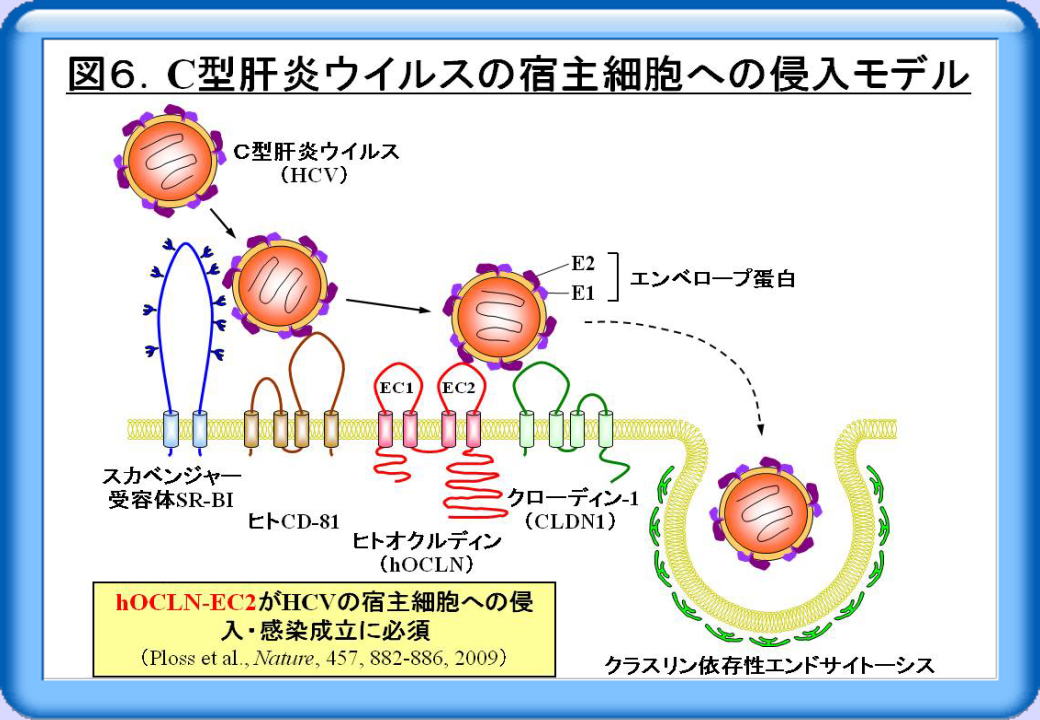
C型肝炎ウィルスは、クラスリン依存性のエンドサイトーシスによって宿主細胞(ヒト肝細胞)に侵入します。この際、スカベンジャー受容体であるSR-BI、ヒトCD-81、Cldn1といった少なくとも3種類の受容体が必要であることが分かっていましたが、これらのみでは感染は成立しません。興味深いことに2009年2月、Rockefeller大学のライスらのグループは、ヒトoccludinの第二細胞外ドメイン (hOCLN-EC2)がC型肝炎ウィルスの宿主細胞への侵入に必須であること、これら四つの受容体があれば線維芽細胞であっても感染が成立することを明らかにしました(図6)。
さらに細胞接着分子や極性分子の中には、癌抑制遺伝子産物として働くものや、癌遺伝子産物の標的となるものがあり、細胞の接着・極性の破綻はがん化の一因と考えられます(図7)。また細胞の接着・極性の破綻によって、がん細胞では増殖因子膜受容体が細胞膜上全体に分布してリガンドと常に会合できる状態にあることが、細胞増殖の異常亢進をきたす一つの要因になるとも考えられます。

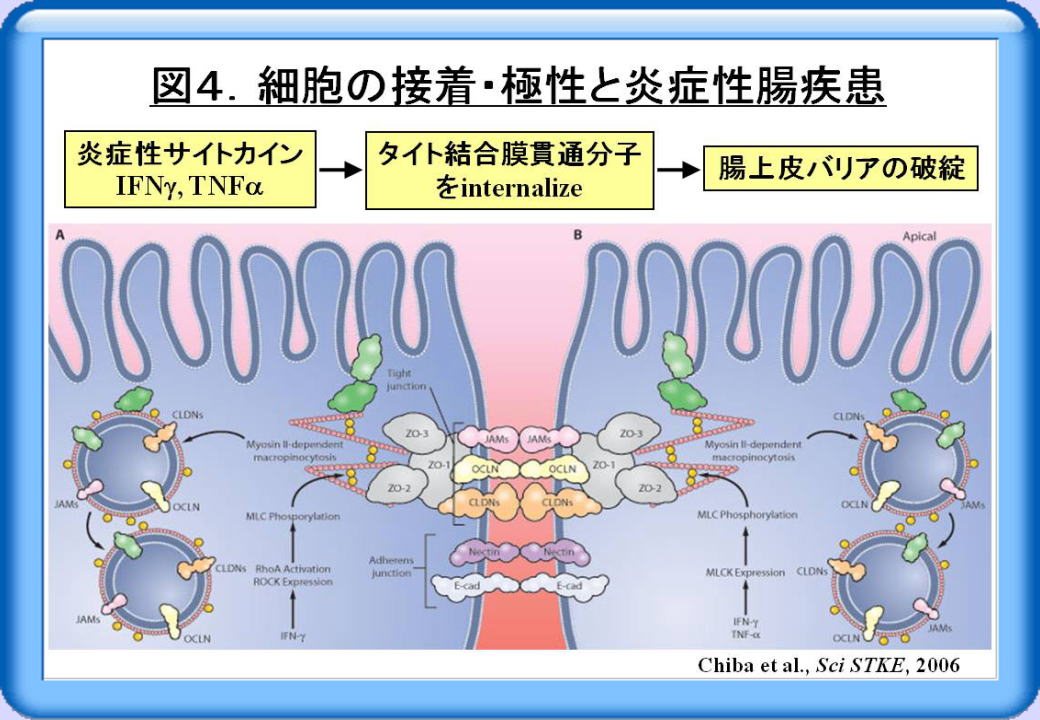
例えば炎症性腸疾患の病態には、炎症性サイトカインであるIFNγ (インタフェロンγ)やTNFα
(腫瘍壊死因子α)が深く関わり、抗TNFα抗体は治療にも用いられています。しかしながら、これらの炎症性サイトカインがどのようにして腸上皮バリアを破綻させるかは明らかでありませんでした。そのような背景の中で2005年末に、IFNγやTNFαが、タイト結合膜貫通分子を細胞内に引き込むこと(エンドサイトーシス)によって、腸上皮バリアを破綻させることが報告されました(図4)。
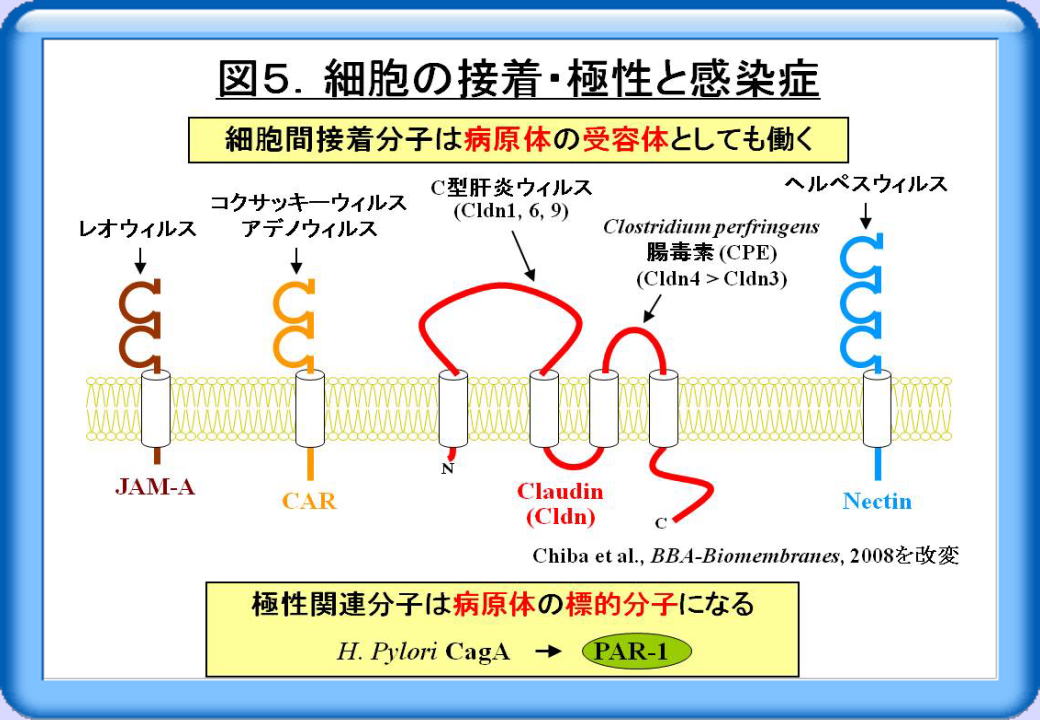
細胞の接着・極性は、感染症とも関連します。例えば、細胞間接着分子JAM-A, CAR, Cldn4, nectinは各々、レオウィルス、コクサッキーウィルス・アデノウィルス、食中毒菌であるClostridium perfringensの腸毒素 (CPE)、ヘルペスウィルスの受容体としても働きます(図5)。また2007年初頭にはCldn1, Cldn6, Cldn9が、C型肝炎ウィルスの受容体として働くことが明らかになりました。さらに極性関連分子PAR-1が、ヘリコバクター・ピロリ菌作り出すCagAと呼ばれるタンパク質の標的分子になること等が分かっています。
1.はじめに
研究においては、まず背景・目的・仮説を明確にすることが大切で、研究の意義を理解することは大きなモチベーションになると考えます。次に、最も適切な材料と方法を選択・構築すること、結果をありのままに捉えること、仮説に捉われずに真実を掴み取ることが重要と思っています。最後に、結果から最も適切な結論を導き出して成果を発表することが研究ではないでしょうか。また研究の過程には、自由な発想と独創性が何よりも肝要な生命線であると考えています。そして、研究の楽しさと面白さを実感してもらえたらと思っています。
私たち基礎病理学講座では、①細胞の接着・極性とヒト疾患、②核内受容体、③幹細胞を主要な研究テーマとしています。これらの研究の背景に触れた後に、これまでの代表的な研究成果、これから目指す研究について紹介させていただきます。
2) 核内受容体による上皮極性と細胞間接着の誘導
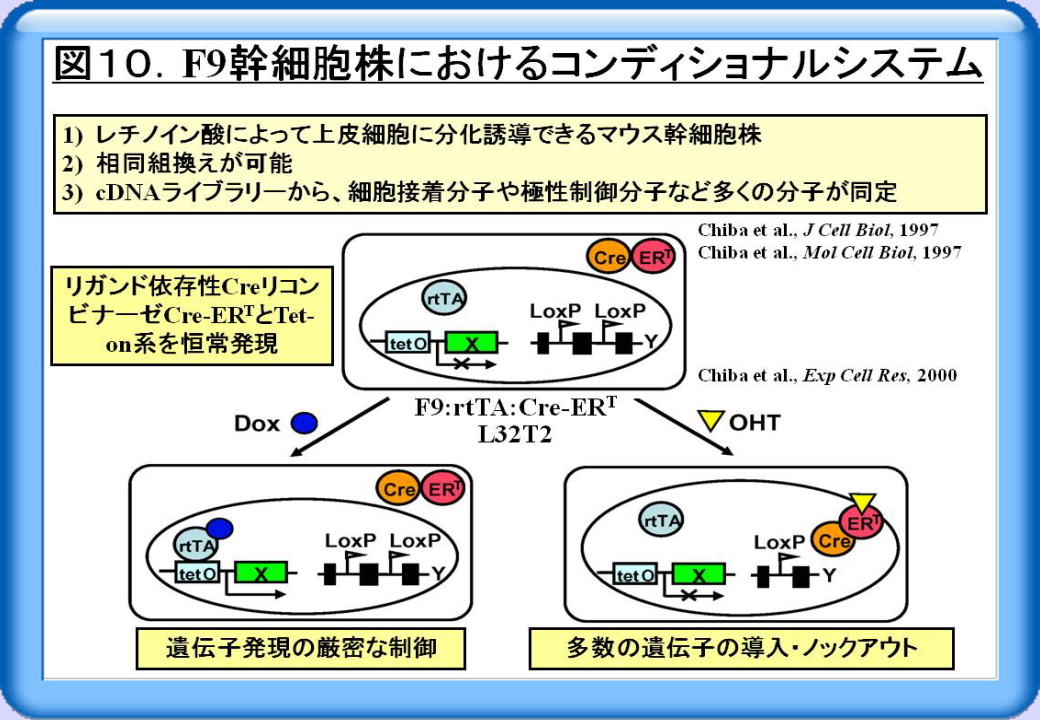
細胞の接着・極性の制御機構や核内受容体の機能を明らかにするために、当教室ではマウスF9幹細胞を一つの材料として用いています。この細胞株はレチノイン酸によって上皮細胞に分化誘導できる幹細胞で、相同組換えが可能です。またF9細胞のcDNAライブラリーから、細胞接着分子や極性制御分子を含む多くの遺伝子が同定されてきた歴史的経緯があり、この細胞を選択しました。
ところで私は、シャンボン研でのポスドク時代に、リガンド依存性CreリコンビナーゼCre-ERTの開発に参画しました。Cre-ERTは、Creリコンビナーゼとヒトエストロゲン受容体リガンド結合領域の変異体との融合タンパク質です。Cre-ERTは内因性のエストラジオール(E2)に非依存性で、合成エストロゲン製剤である4-水酸化タモキシフェン(OHT)投与によって活性化されます。細胞特異的プロモーターによってCre-ERT遺伝子の発現を制御したトランスジェニックを用いると、マウス個体レベルで遺伝子変異の部位と時期と特定することができ、この系はコンディショナルノックアウトマウスのスタンダードとなっています。
そこでF9幹細胞において、Tet-on系とCre-ERTを恒常発現させ、F9:rtTA:Cre-ERT L32T2という細胞株を樹立しました(図 10)。本細胞株では、テトラサイクリン誘導体であるドキシサイクリン(Dox)で処置することによって、遺伝子発現の厳密な制 御が可能です。またOHTで処置すると、多数の遺伝子を導入・ノックアウトすることができます。例えばこの細胞株では、loxP 配列で挟まれた耐性遺伝子発現ベクター切り取ることができますので、同じ耐性遺伝子発現ベクターを何度でも使って多数 の遺伝子を導入できます。またRNAi法による遺伝子ノックダウンもワークし、種々の遺伝子操作が可能な理想的なコンディ ショナルシステムです。
2.細胞の接着・極性とヒト疾患/核内受容体
1) 研究の背景
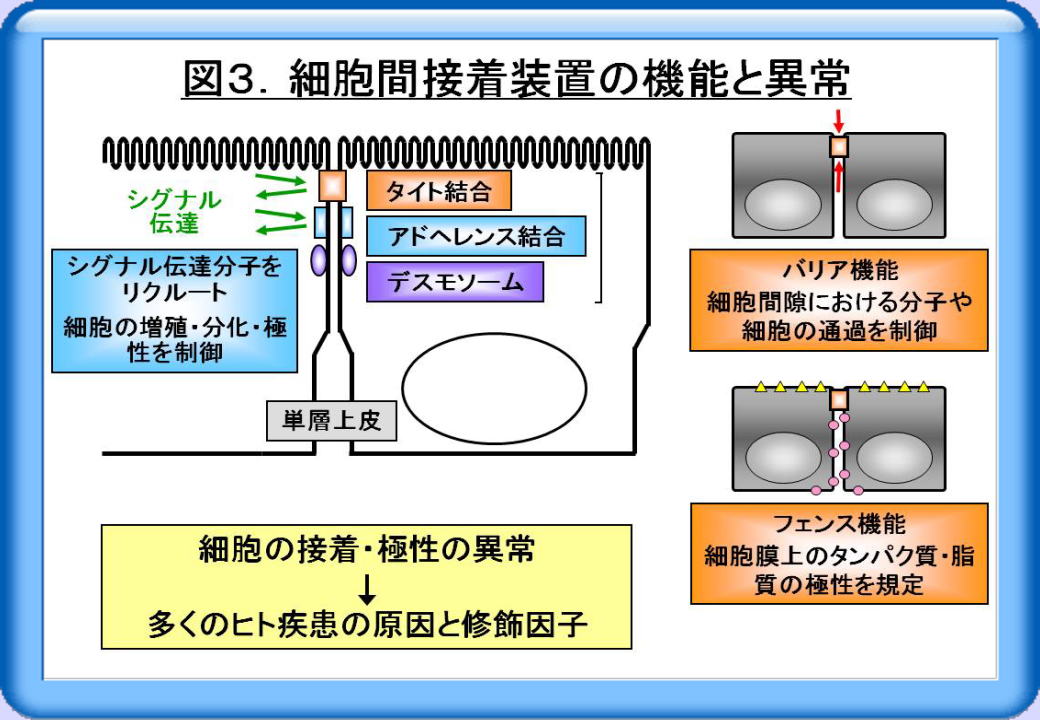
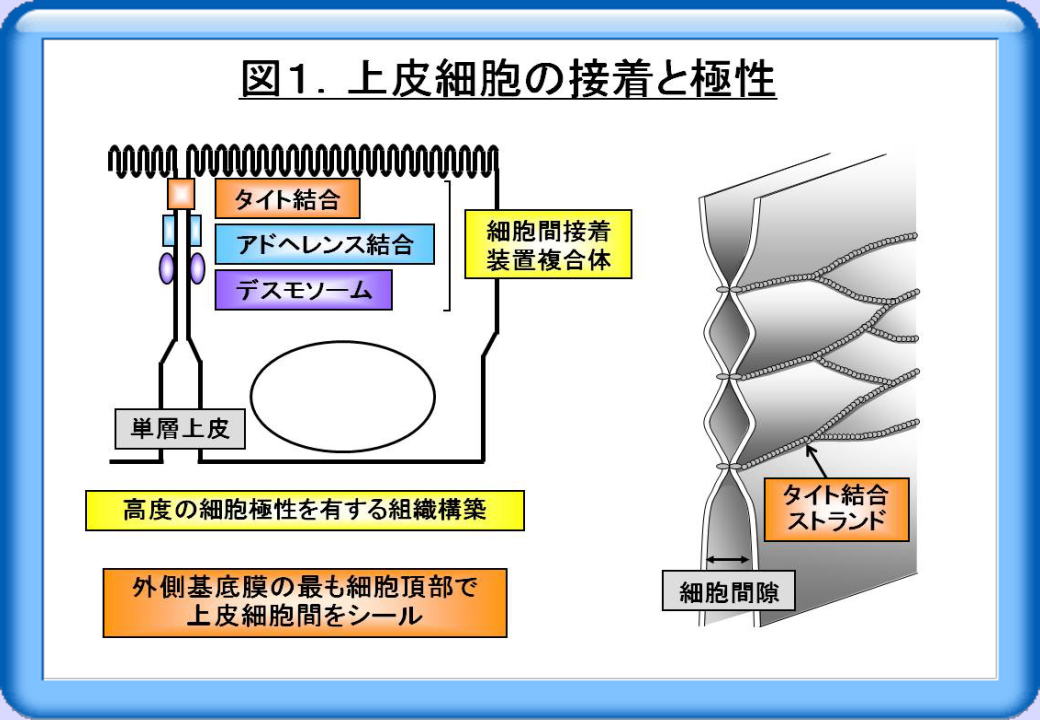
ヒトをはじめとする多細胞生物の生命現象には、極性を持った細胞の存在や細胞接着が不可欠です。例えば体表や体腔を覆う上皮細胞では、タイト結合・アドへレンス結合・デスモソームからなる細胞間接着装置がよく発達しており、 高度の細胞極性を有する組織構築が形成・維持されています(図1)。このうちタイト結合は、外側基底膜の最も細胞頂部で上皮細胞間をシールし、タイト結合ストランドと呼ばれる膜内粒子の連続した紐状の超微構造を形成しています。
研究内容
私たちはこのようなコンディショナルシステムを用いて、核内受容体スーパーファミリーに属するレチノイド受容体とHNFα(hepatocyte nuclear
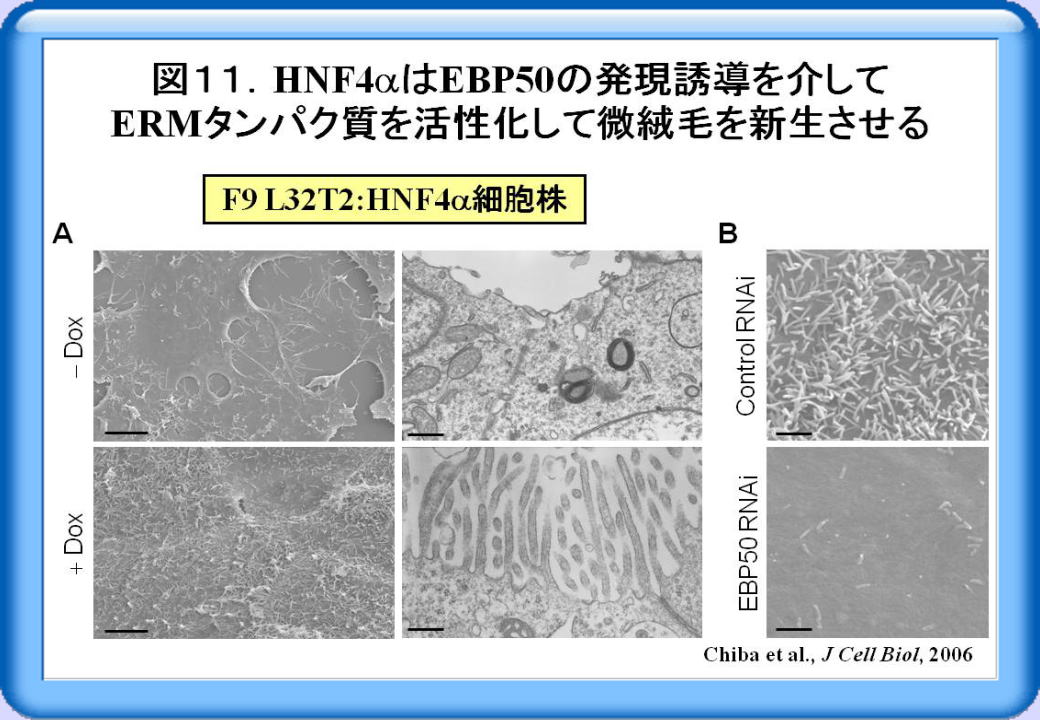
factor α)が上皮極性と細胞間接着の形成を誘導することを明らかにしました。図11Aは、DoxによってHNFαの発現を誘導できるF9細胞株(F9 L32T2:
HNFα)の超微構造を走査電顕と透過電顕で観察したものです。上段の未分化F9幹細胞では、糸状突起様構造がみられますが、微絨毛は殆んど認められません。一方DoxによってHNFαの発現を誘導した細胞では、微絨毛の数と長さが著明に増加しており、HNFαが微絨毛を著しく誘導することを発見しました。また微絨毛関連分子のうち、EBP50(ezrin/radixin/moesin
[ERM]-binding phosphoprotein 50)の発現が、HNFαによって著しく誘導されることを見出しました。さらにEBP50の発現をノックダウンすると微絨毛形成が著明に抑制された(図11B)ことなどから、HNFαはEBP50の発現誘導を介してERMタンパク質を活性化して微絨毛を新生させることを突き止めました。
3) 腸上皮細胞間隙における新たなカルシウム吸収機構
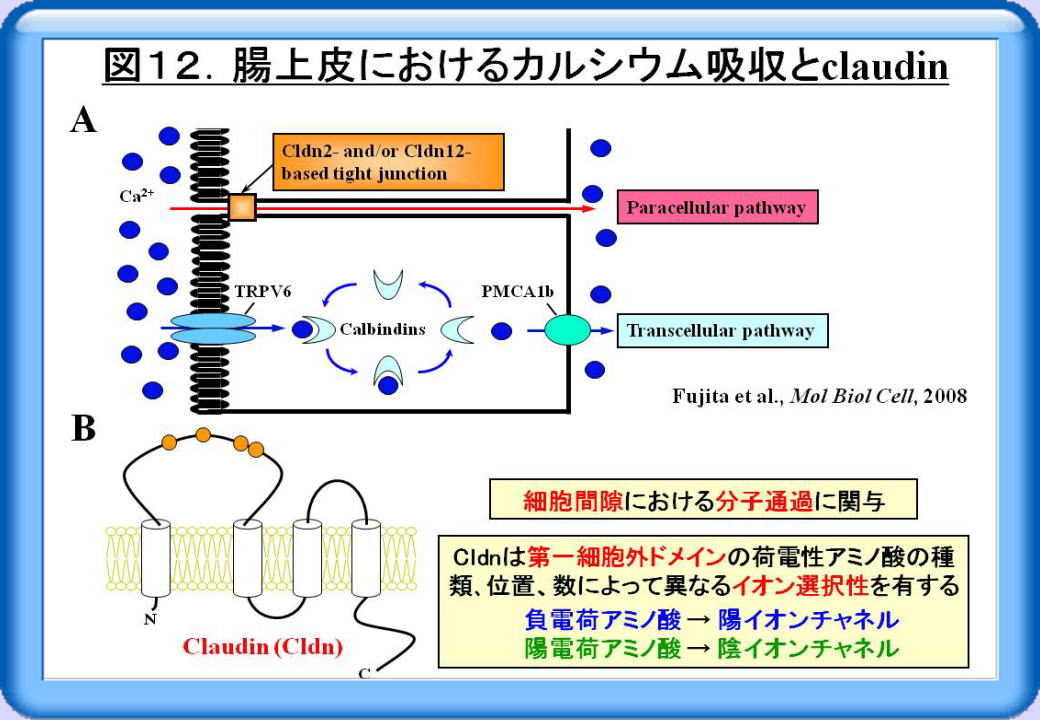
カルシウムイオン (Ca2+) は腸上皮細胞から吸収され、その経路には細胞内を通るtranscellular pathwayと細胞間隙を通るparacellular pathwayが知られています(図12A)。また活性型ビタミンD3は、transcellular pathwayによるCa2+吸収を亢進させる最も重要なホルモンの一つです。しかし、paracellular pathwayを介したCa2+吸収に関わる分子や、そのビタミンD3依存性は不明でした。
細胞間隙における分子通過、すなわちタイト結合のバリア機能にとって最も重要な分子が、前述したclaudin(Cldn)です。またCldnは、第一細胞外ドメインの荷電性アミノ酸の種類、位置、数によって異なるイオン選択性を有することが明らかになってきました(図12B)。すなわち、負電荷アミノ酸が多いと陽イオンチャネルに対する細胞間隙チャネル、陽電荷アミノ酸が多いと陰イオンチャネルに対する細胞間隙チャネルとして働くことが示唆されています。
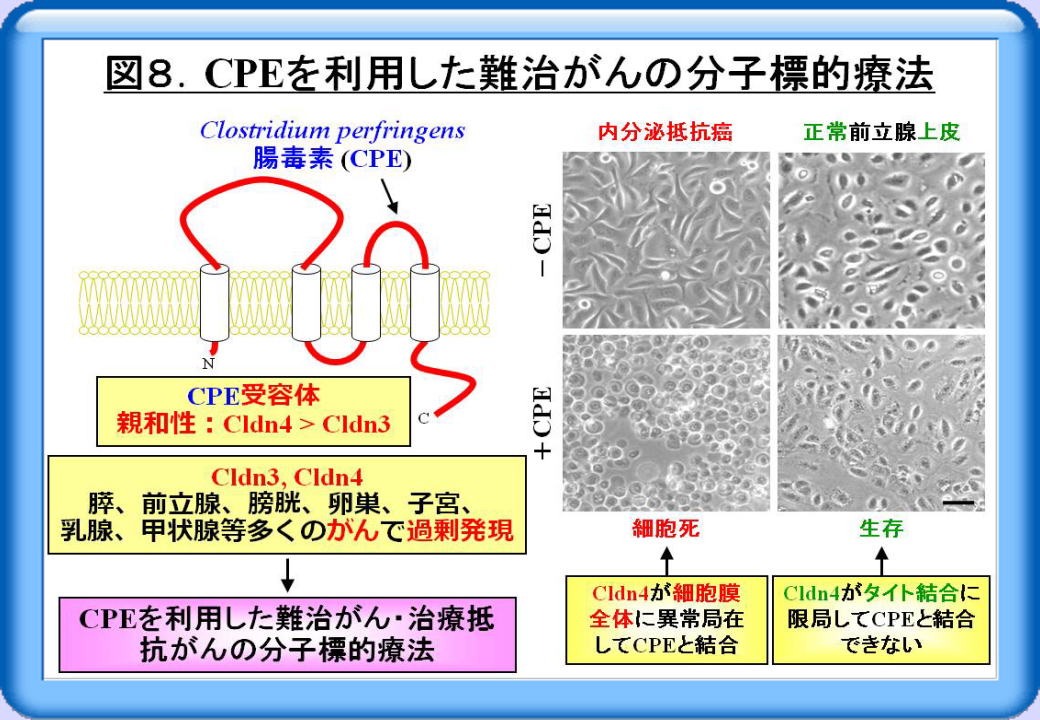
前述した通りCldn4は、CPEの受容体として働きますが、多くのがんで過剰発現しています。そこで、CPEを利用した難治がん・治療抵抗がんの分子標的療法が期待を集めています。例えば、CPEで処置した内分泌抵抗前立腺がん細胞は細胞死に至りますが、正常前立腺上皮細胞はCPE存在下でも生存できるという結果を得ています(図8)。詳しいデータは示ししませんが、Cldn4が細胞膜全体に異常局在しているがん細胞ではCPEと結合でき、Cldn4がタイト結合に限局している正常細胞ではCPEと結合できないと考えられました。したがってCPEによって、正常細胞に影響を及ぼさずにがん細胞だけを狙い撃ちできる可能性が示唆されました。
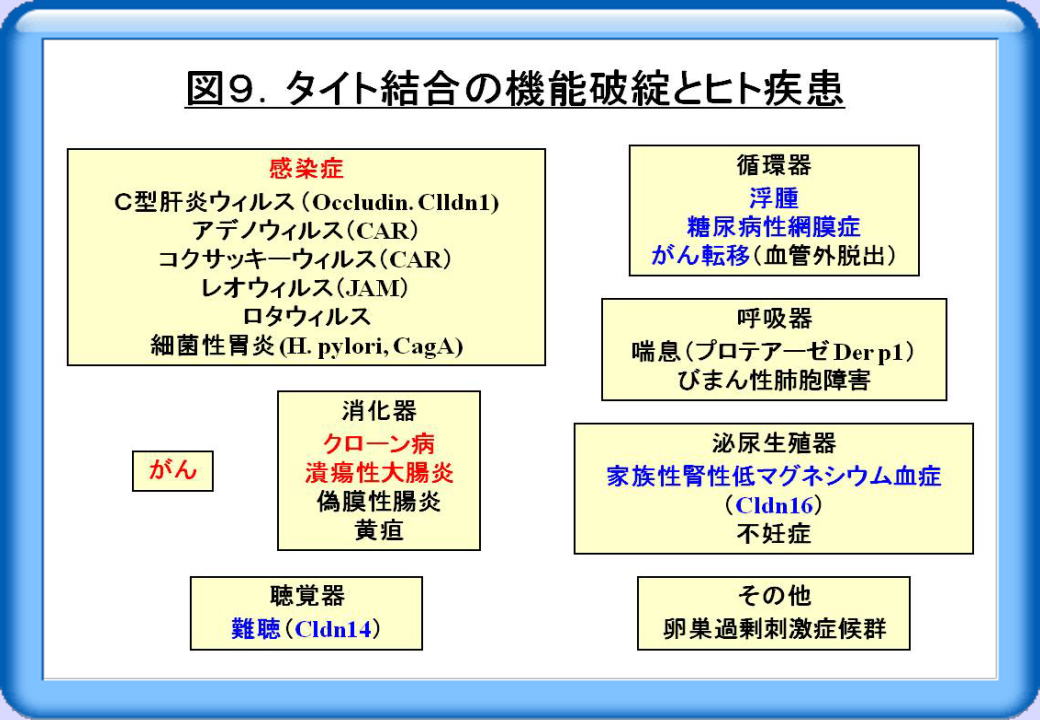
これまで細胞の接着・極性と炎症性腸疾患、感染症、がんについて述べましたが、タイト結合の機能破綻とヒト疾患に限っても、その他多くの報告があります(図9)。
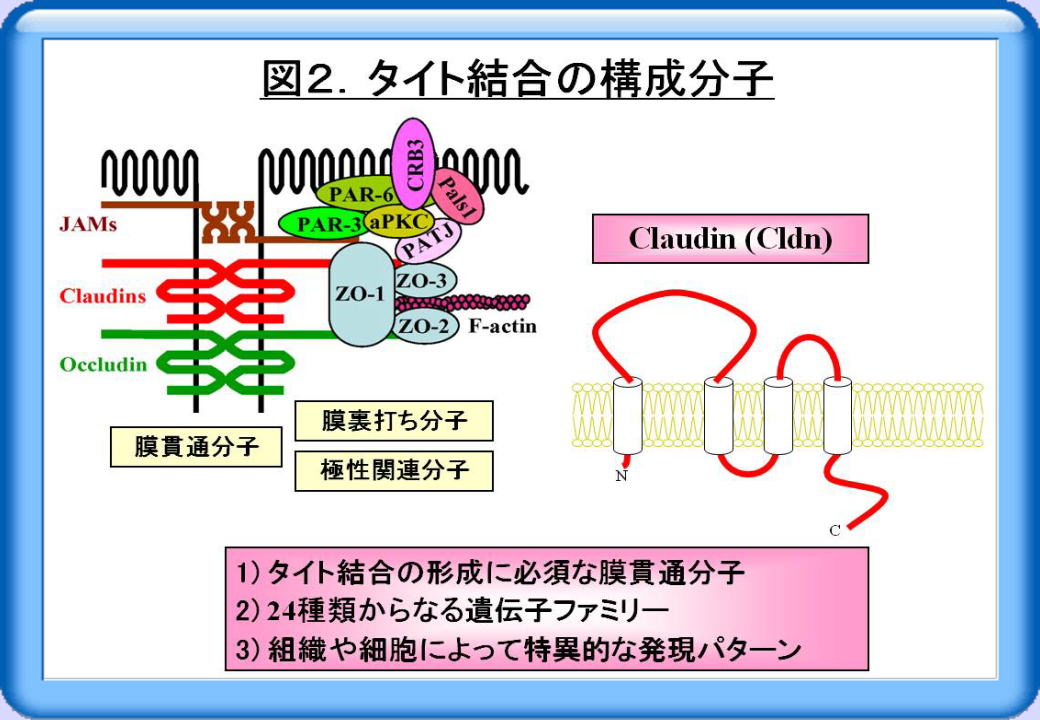
タイト結合の構成分子としては、occludin, claudin, JAM (junctional adhesion molecule)といった膜貫通分子と、多くの膜裏打ち分子や極性関連分子が知られています(図2)。このうちclaudin (Cldn)は、タイト結合の形成に必須の膜貫通分子です。またCldnは、24種類からなる遺伝子ファミリーを構成し、組織や細胞によって特異的な発現パターンを示します。
これら細胞間接着装置は機械的な結合に関わるだけでなく、シグナル伝達分子をリクルートして、細胞の増殖・分化・極性を制御しています(図3)。さらにタイト結合は、細胞間隙における分子や細胞の通過を制御するバリア機能や、細胞膜上のタンパク質・脂質の極性を規定するフェンス機能を担っています。したがって細胞の接着・極性の異常は、多くのヒト疾患の原因と修飾因子となります。

そこで、「腸上皮に発現し且つ第一細胞外ドメインに負電荷アミノ酸を多く有するCldn、すなわちCldn 2, 7, 12, 15のいずれかが、Ca2+のparacellular channelとして働き、Ca2+の細胞間隙通過を制御する」、「これらのCldnsの発現は活性型ビタミンD3によって誘導される」という仮説を立てました。
ビタミンD受容体(VDR)ノックアウトマウスやビタミンD依存性腸上皮細胞株Caco-2を用いて検討した結果、活性型ビタミンD3がVDRを介して、腸上皮におけるCldn2とCldn12の発現を誘導することを見出しました。また、Cldn2とCldn12からなるタイト結合は、腸上皮細胞間でCa2+のparacellular channelを形成し、Ca2+の細胞間隙通過を制御することを突き止めました(図12A)。カルシウムイオンのparacellular channelを同定した本研究を基盤として、カルシウムを効率よく吸収・通過させることが可能になれば、骨粗鬆症やカルシウム代謝異常に対する新たな予防・治療戦略になることが期待されます。