
貝淵 弘三(名古屋大学)

「リン酸化シグナルに基づいた報酬計神経回路の操作技術開発」
快情動行動や報酬学習の神経基盤として、側坐核と前頭前皮質、扁桃体や海馬などの関連領域で形成される報酬系神経回路が重要な役割を果たしています。側坐核にはドーパミンD1受容体(D1R)を発現する中型有棘神経細胞、ドーパミンD2受容体(D2R)を発現する中型有棘神経細胞、GABAおよびアセチルコリン作動性介在神経細胞が存在し、約90%は中型有棘神経細胞で占められています。ドーパミン、セロトニン、アセチルコリン、アデノシンなどはニューロモジュレーターと呼ばれ、中型有棘神経細胞に存在するG蛋白質共役型受容体(GPCR)を介して細胞内のリン酸化シグナルを調節し、神経細胞の興奮性や可塑性を通じて側坐核の働きを制御します。
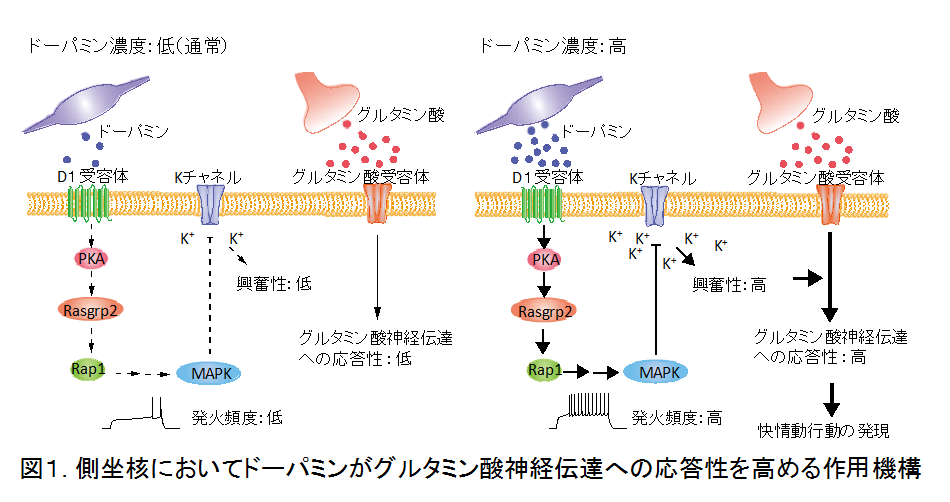
快情動行動が発現する際、ドーパミンによって側坐核のD1R-中型有棘神経細胞が興奮性の低い状態から高い状態へと「シフト」することで神経回路が作動し快情動行動が発現します。一方、報酬学習の際にはドーパミンによって長期増強や遺伝子発現などの長期的な現象が引き起こされ、それによって神経回路の活動が促進されることで報酬学習が成立すると考えられています。しかしながら、側坐核神経回路の活動性のシフトが調節される詳細な機構はわかっていません。
これまでに私たちは、新たに開発したリン酸化プロテオミクス法により、ドーパミンによって引き起こされる中型有棘神経細胞の細胞内シグナルを網羅的に解析し、ドーパミンがD1R-PKA-Rap1-MAPK経路を活性化させることで、D1R-中型有棘神経細胞を興奮性の低い状態から高い状態へとシフトさせ、グルタミン酸神経伝達への応答性を高めていることを見出しています(図1)。これはドーパミンが快情動行動を生み出す分子メカニズムを初めて明らかにしたものであると考えられます。
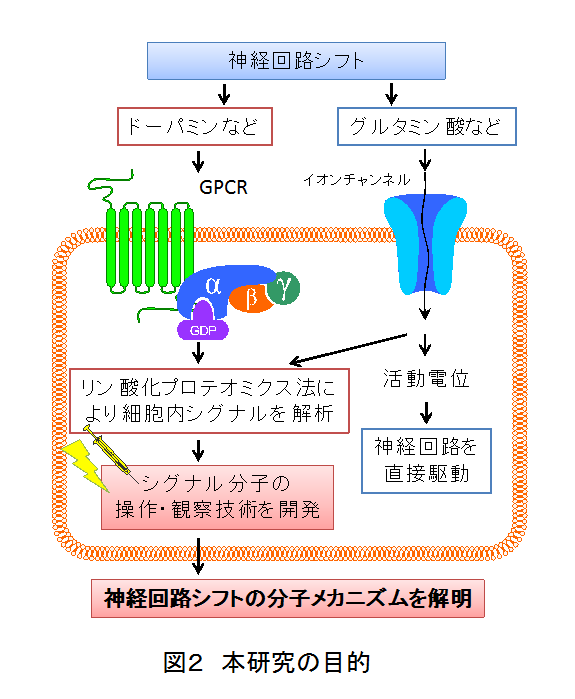
本研究では、報酬系神経回路の機能シフトに関連するリン酸化シグナルを網羅的に解析し、それに基づいた新しい報酬系神経回路の操作技術を開発します。さらに、神経細胞の興奮性や可塑性が回路シフトを制御する機構について明らかにします。
最近の主要論文
1. Amano M, Nishioka T, Yura Y, Kaibuchi K. Identification of Protein Kinase Substrates by the Kinase-Interacting Substrate Screening (KISS) Approach. Curr Protoc Cell Biol. 72:14.16.1-14.16.12. (2016)
2. Nagai T, Yoshimoto J, Kannon T, Kuroda K, Kaibuchi K. Phosphorylation Signals in Striatal Medium Spiny Neurons. Trends Pharmacol Sci. 37(10):858-71. (2016)
3. Nagai T, Nakamuta S, Kuroda K, Nakauchi S, Nishioka T, Takano T, Zhang X, Tsuboi D, Funahashi Y, Nakano T, Yoshimoto J, Kobayashi K, Uchigashima M, Watanabe M, Miura M, Nishi A, Kobayashi K, Yamada K, Amano M, Kaibuchi K. Phosphoproteomics of the Dopamine Pathway Enables Discovery of Rap1 Activation as a Reward Signal In Vivo. Neuron. 89(3):550-65. (2016)
4. Xu C, Funahashi Y, Watanabe T, Takano T, Nakamuta S, Namba T, Kaibuchi K. Radial Glial Cell-Neuron Interaction Directs Axon Formation at the Opposite Side of the Neuron from the Contact Site. J Neurosci. 35(43):14517-32. (2015)
投稿日:2017年05月07日